丂丂俁丏丂俢俶俙揮幨偺儊僇僯僘儉乮俀乯
丂丂乮係乯丂俢俶俙偺帺屓暋惢 偲 撍慠曄堎丗
丂丂嵶朎暘楐偺偲偒偺俢俶俙偺帺屓暋惢偼丄俀杮偐傜侾杮偵暘偐傟偨偦傟偧傟偺曅懁偺傒偺暋惢偱偁傝丄乽敿曐懚揑暋惢乿偱偁傞丅 峺慺偺僿儕僇乕僛偑嵔傪姫偒栠偟夝偒傎偖偟丄億儕儊儔乕僛偑暥帤傪撉傒庢傝丄曅懁偺傒偺嵔傪怴偟偔崌惉偡傞丅俢俶俙偑偦傟偧傟 侾丗侾懳墳偡傞墫婎楍偺俀杮嵔偱偱偒偰偄傞偺偱丄偦傟偑僘儃儞偺僠儍僢僋偺傛偆偵侾侽墫婎傎偳奐偒丄俀杮嵔摨帪偵帺慠揑偵暋惢偑惉偝傟偰偄偔丅
丂丂傑偨丄恊偐傜偺堚揱偼丄僸僩偺応崌丄愼怓懱偼丄俀俀懳偺忢愼怓懱偲 侾懳偺惈愼怓懱偺 寁係俇杮偱偁傝丄晝偲曣偐傜侾杮偢偮揱偊傜傟傞丅
丂丂岎攝偵傛偭偰偼堎側傞庬乮偟傘乯摨巑乮偨偲偊偽丄僸僩偲僒儖丄僀僰偲僱僐側偳乯偱偼寛偟偰巕懛傪嶌傞偙偲偑偱偒側偄丅庬偼忢偵撈棫偟偰偄傞丅 偨偩偟丄庬撪偱偼丄偄偔偮偐偺岎攝庬偑敪惗偟偆傞丅乮僂僔壢丗 岎攝壜擻側偺偼丄傾僲傾丄僶僀僜儞丄僈僂傾丄傾僕傾僗僀僊儏僂丄傾僼儕僇僗僀僊儏僂丄儎僋側偳丅 僀僰壢(僕儍僢僇儖丄僆僆僇儈丄僐儓乕僥側偳乯丅 僂儅偲儘僶偼儔僶傪惗傓偑丄梇儔僶偵偼惗怋擻椡偑柍偔丄帗儔僶偼幚幙僂儅偱偁傞丅丂丒丒丒丂岎攝偺尷奅偵嬤偄乭嶨庬乭偺巕偼丄惗怋擻椡偑柍偔愨柵偡傞偐丄偁偭偰傕偦偺巕懛偼偳偪傜偐偺恊偺庬偵栠偭偰偟傑偆乯
丂丂丂乽偮偄偱恄偼丄乽抧偼丄偦偺庬椶偵偟偨偑偭偰丄惗偒暔丄壠抺傗丄偼偆傕偺丄偦偺庬椶偵偟偨偑偭偰栰偺廱傪惗偤傛丅乿偲嬄偣傜傟偨丅偡傞偲偦偺傛偆偵側偭偨丅恄偼丄偦偺庬椶偵偟偨偑偭偰栰偺廱丄偦偺庬椶偵偟偨偑偭偰壠抺丄偦偺庬椶偵偟偨偑偭偰抧偺偡傋偰偺偼偆傕偺傪憿傜傟偨丅恄偼尒偰丄偦傟傪傛偟偲偝傟偨丅乿乮憂悽婰侾丗俀係丄俀俆乯
丂丂丂丂丂丂丂丂丂丂丒丒丒丒丒丒丂乭庬乭偵廬偭偰偺乽摿暿憂憿乿丅偡傋偰偺庬偼尰嵼傑偱傕姰慡偵撈棫偟偰偄傞丅恑壔榑偺乭宯摑庽乭偼懚嵼偟摼側偄丅
丂丂偝偰丄俢俶俙暘巕偑帺屓暋惢偡傞嵺丄偁傞乽妋棪乿偱丄傑傟偵娫堘偄傪惗偠傞丅乮仏 尰嵼偼丄偙傟傪乽撍慠曄堎乿偲屇傇乯
丂丂墫婎偺 抲姺傛傝傕丄寚幐丄憓擖乮摿偵丄墫婎偺孞傝曉偟攝楍乮俠俧俠俧丒丒丒側偳乯偺屄悢傪曄壔偝偣傞曄堎乯偑丄傛傝崅偄妋棪偱婲偙傝傗偡偄丅偦偺偨傔丄偙偺孞傝曉偟晹暘亖乭儅僀僋儘僒僥儔僀僩懡宆乭偲屇偽傟傞椞堟 偱丄乭俢俶俙巜栦乭偲偟偰 俢俶俙偺徻嵶側屄懱嵎傪挷傋傞偺偵梡偄傜傟傞丅
丂丂偙偺丄俢俶俙暋惢偵偍偗傞撍慠曄堎偑巆偭偰偄傞売強偼丄埑搢揑偵僀儞僩儘儞丄偡側傢偪丄慡俢俶俙偺俋俆亾埲忋傪愯傔傞乭偑傜偔偨堚揱巕乭偺晹暘偱偁傞丅
丂丂側偤側傜丄堚揱忣曬傪偵側偆僄僋僜儞偺晹暘偵曄堎偑婲偙傞偲丄偦偺曄堎偺傎偲傫偳偼惗懚偵晄棙側撍慠曄堎偱偁傝丄偦偺嵶朎偑丄庴惛棏偺応崌偼 偦偺屄懱傑偱傕偑丄懍傗偐偵巰柵偡傞偐丄抳柦揑側塭嬁傪庴偗偰偟傑偆偐傜偱偁傞丅 偟偨偑偭偰丄乭撍慠曄堎乭偲尵偭偰傕丄傔偭偨側偙偲偱偼乭庬乭偵偼帠幚忋傎偲傫偳壗偺曄壔傕傕偨傜偝側偄偺偱偁傞丅 惗懚偵桳棙偱傕晄棙偱傕側偄曄堎 亖 乭拞棫撍慠曄堎愢乭乮栘懞丒1968乯偼偙偺僀儞僩儘儞偺晹暘偱惉棫偡傞偲尵偆偙偲偑偱偒傞偑丄尰傟偨屄懱偺宍幙偼 幚嵺偵偼壗傕曄傢偭偰偍傜偢丄偄傢備傞乭恑壔乭偱偼側偔丄側傫偲偐乭尰忬堐帩乭偟偰偄傞丄偲尵偭偨曽偑惓偟偄丅
丂丂仏丂偨偲偊偽丄乭惗偒偰偄傞壔愇乭偲尵傢傟傞乭僔乕儔僇儞僗乮攛嫑偺堦庬偱 僄儔偲丄攛偺婡擻傪帩偮晜偒戃偺椉曽傪帩偮乯乭偼丄嫑椶偲椉惗椶偲偺拞娫庬偲峫偊傜傟偨帪婜傕偁偭偨偑丄敿悽婭慜儅僟僈僗僇儖搰壂偱曔妉偝傟偨傕偺傪挷嵏偟偨寢壥丄俈侽侽侽擭慜偲偄傢傟傞壔愇偲壗堦偮乭恑壔乭傗乭曄壔乭偑擣傔傜傟側偐偭偨丅傑偨丄僀儞僪僱僔傾偺僐儌僪僩僇僎乮懱挿俁亅俉倣乯傕丄偦偺壔愇偲壗偺曄壔傕擣傔傜傟側偄丅
丂丂仏仏丂儊僉僔僐偺傾僇儞僶儘攷暔娰偵揥帵偝傟偰偄傞懡偔偺乭嫲棾搚嬼乭偼丄乭擬儖儈僱僙儞僗朄乮俿俴朄乯乭偱栺係俆侽侽擭慜乮岆嵎俆乣侾侽亾乯偲弌偨偑丄偙傟偼乽僲傾偺峖悈乿偑婲偙偭偨帪婜乮俛丏俠丏俀侽侽侽擭慜屻乯偲傎傏堦抳偡傞丅丂偙傟偼丄戝峖悈屻丄偟偽傜偔偺婜娫偼丄嫲棾偑乮偐側傝彫偝偔側偭偨偑乯惗偒偰偄偨丄偲偄偆偙偲偱偁傞丅乮偦偺彫偝側嫲棾偵恖娫偑忔偭偰偄傞傕偺傕偁傞乯丂偙傟偼丄峖悈屻偺娐嫬偺寖曄乮婥埑丗 俀乣俁婥埑 仺 侾婥埑丄嬻婥偺巁慺擹搙丗 俁侽亾 仺 俀侾亾丄婥壏偺姦椻壔丄怉惗偺曄壔丄側偳乯偵傛偭偰丄寣偺弰傝偺埆偄 嫲棾偼戝偒偔惉挿偱偒側偐偭偨偨傔偲巚傢傟傞丅丂偙偺応崌偵偼丄乭帺慠搼懣乭偲偄偆偙偲偑偱偒傞丅
丂丂栺係侽侽侽擭慜偺乽僲傾偺峖悈乿偵傛偭偰丄旕忢偵懡偔偺惗暔偑乭帺慠搼懣乭偝傟丄偦偺巆奫偼壔愇偲側偭偰偄傞丅乮弖娫揑偵搚嵒偵暵偠崬傔傜傟側偄尷傝寛偟偰壔愇偲側傜側偄乯
丂
丂丂乮俆乯丂僸僩俢俶俙偺捛愓挷嵏丗
丂丂曄堎偺宱楌偑巆傞 僀儞僩儘儞偺撍慠曄堎売強傪挷傋傞偙偲偵傛偭偰丄庬偑偳偺傛偆側宱堒傪偨偳偭偰曄壔偟偰偒偨偐傪抦傞偙偲偑偱偒傞丅
丂丂嘆丂儈僩僐儞僪儕傾俢俶俙丗
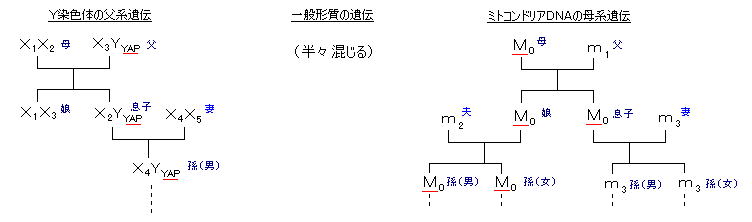
丂丂嵶朎撪彫婍姱偺堦偮偱偁傞 儈僩僐儞僪儕傾乮嵶朎撪屇媧傪偮偐偝偳傞彫婍姱乯偵傕丄彫偝偄側偑傜撈帺偺俢俶俙偑偁傞丅儈僩僐儞僪儕傾俢俶俙偼丄墫婎悢 栺侾俇俆侽侽屄偱偁傝丄曣惈堚揱偡傞丅乮惛巕偵傕儈僩僐儞僪儕傾俢俶俙偑懚嵼偡傞偑丄庴惛棏偺拞偱攋夡偝傟傞乯丂妀堚揱巕偺堦斒宍幙偼丄晝偲曣偐傜敿暘偢偮庴偗宲偑傟傞偨傔丄堚揱嫍棧偺憡娭恾偑暘偐傞偺傒偱偁傞偺偵懳偟丄儈僩僐儞僪儕傾俢俶俙偼曣偐傜偺傒揱偊傜傟傞偺偱丄抝宯傪柍帇偟偨彈宯偩偗偺乽宯恾乿傪嶌傞偙偲偑偱偒丄偙偺堚揱巕慡懱偺嫟捠慶愭偼 偁傞堦恖偺彈惈偺儈僩僐儞僪儕傾乮乭儈僩僐儞僪儕傾丒僀僽乭乯偵偨偳傝拝偔偙偲偵側傞丅
丂丂偟偐傕偙偺宯恾偼丄俢俶俙偺曄堎妋棪偑堦掕偱偁傞偲偡傞偲丄偦偺暘婒偐傜偺帪娫偑悇掕偱偒傞丅偡側傢偪丄偳偺帪婜偵偳偺傛偆側恖庬偑暘婒偟偨偐傪丄懠偺峫屆妛揑僨乕僞乮14俠朄側偳乯偲嫟偵悇掕偟丄捛愓偡傞偙偲偑偱偒傞丅乮仏丂儈僩僐儞僪儕傾俢俶俙偺曽偑丄倄愼怓懱俢俶俙偺僀儞僩儘儞晹暘傛傝傕曄堎懍搙偑懍偄偺偱丄偨偲偊偽傾僕傾抧堟側偳偺嫹偄椞堟偺尷傜傟偨帪娫撪偵偍偗傞曄壔傪挷傋傞偺偵梡偄傜傟傞丅乯
丂丂嘇丂倄愼怓懱俢俶俙丗
丂丂妀俢俶俙偼丄栺俇侽壄屄偺墫婎乮儈僩僐儞僪儕傾偺係侽枩攞乯偱偁傞偑丄惈愼怓懱乮抝丗 倃倄丄彈丗 倃倃乯偺偆偪偺 倄愼怓懱偺俢俶俙偼丄扨弮偵晝偐傜懅巕傊偺傒揱傢傞偺偱丄抝宯偺堚揱巕宯恾傪嶌傞偙偲偑偱偒丄儈僩僐儞僪儕傾俢俶俙偲摨條偵丄暘婒帪婜傗偳偺傛偆偵堏摦偟偨偐傪悇掕偡傞偙偲偑偱偒傞丅 偦傟偧傟偺僌儖乕僾偺壠宯偱乭抝宯乭偑庣傜傟偰偄傞側傜偽丄惓妋側捛愓寢壥偲側傞丅
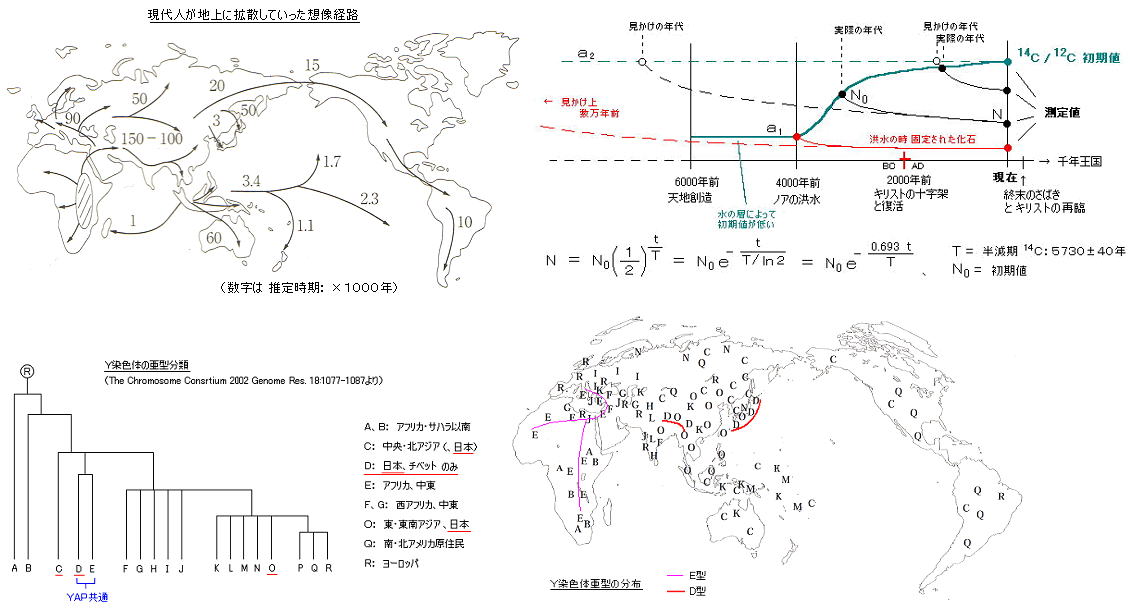
丂丂摿偵丄乭倄俙俹乭乮辕踢乯偲屇偽傟傞曄堎売強偼 暘椶忋 俢丄俤 宯摑偺僌儖乕僾偩偗偑桳偟丄擔杮偺撽暥宯偲拞搶丒傾僼儕僇宯偩偗偵嫟捠偡傞傕偺偲偟偰丄偦偺嬤墢惈偑拲栚偝傟偰偄傞丅
丂丂
丂丂偦傟埲慜乮侾俋俇侽擭戙乯偵峴側傢傟偨丄寣塼拞偺偨傫傁偔幙偺傾儈僲巁攝楍偵傛傞傾僕傾恖庬偺嬤墢惈挷嵏寢壥偱偼丄傾僕傾俇廤抍乮娯崙恖丄儌儞僑儖恖丄杮搚擔杮恖丄僠儀僢僩恖丄傾僀僰恖丄壂撽恖乯偺俢俶俙嬤墢惈偺傒偑妋擣偝傟丄撿晹拞崙恖丄搶撿傾僕傾恖庬偺僌儖乕僾丄傑偨丄杒暷尨廧柉丄僄僗僉儌乕丄撿暷尨廧柉偺僌儖乕僾偲偼丄偦傟偧傟嫍棧傪抲偄偨傕偺偲側偭偨丅偟偐偟丄寣塼拞偺偨傫傁偔幙偺傾儈僲巁攝楍偼丄妀俢俶俙偺僄僋僜儞晹暘亖曄堎偑晹暘揑偵偟偐巆傜側偄晹暘偵傛傞偺偱丄晄姰慡側寢壥偱偁傞丅乮敡偺怓傗丄懱宆丄婄偐偨偪側偳偺庒姳偺曄壔偼丄惗懚偵桳棙偱傕晄棙偱傕側偄応崌偼丄曄堎偟偰傕栤戣側偔堷偒宲偑傟傞偩傠偆乯
丂丂偦偟偰丄偙偺俀侽擭娫偺尋媶偵傛傝丄儈僩僐儞僪儕傾傗 倄愼怓懱偺俢俶俙偺挷嵏偱偼丄宯恾偑嶌傜傟丄暘婒偲堏摦偺帪婜傑偱丄傎傏惓妋偵悇掕偡傞偙偲偑偱偒傞傛偆偵側偭偨丅
丂丂尰嵼偺倄愼怓懱垷宆偺悽奅暘晍偵傛傞偲丄擔杮恖偺栺係侽亾乮撽暥宯丄俢2宯摑丒僴儉宯乯偼丄乮尒偨栚忋偺堦斒宍幙偼傎偲傫偳傾僕傾恖偱偼偁傞偑乯幚偼側傫偲丄拞搶丒傾僼儕僇恖偵嬤偄乮俤宯摑丒僴儉宯乯丅 俢宯摑偲 俤宯摑偼 屆偄帪婜偵暘婒偟偨傕偺偱偁傝丄抝宯偑庣傜傟偰偄傞側傜偽丄撽暥恖偺慶愭偼偐側傝屆偄帪婜偵傾僼儕僇曽柺偐傜傗偭偰棃偨偙偲偵側傞丅
丂丂偙偺挷嵏偝傟偨俢俶俙売強偼丄曄堎偺棜楌偑拤幚偵巆傞僀儞僩儘儞偺晹暘偱偁傝丄倄俙俹偲偄偆栚報傑偱晅偄偰偄傞丅乮妋偐偵丄暥壔揑側傕偺傪彍偄偰傕丄擔杮恖偺惛恄峔憿偼懠偺傾僕傾恖庬偲偳偙偐堘偭偰偄傞丅丂仏丂曣惈堚揱偡傞儈僩僐儞僪儕傾俢俶俙偵傛偭偰傕丄擔杮恖偺拞偵拞搶丒傾僼儕僇宯偑栺侾俆亾懚嵼偡傞丅乯
丂丂壓偺嵍忋恾乮乽俢俶俙偐傜尒偨擔杮恖乿丄嵵摗挊丄偪偔傑怴彂丄2005丄倫俇侽乯偺丄乭尰戙恖偑抧忋偵奼嶶偟偰偄偭偨憐憸宱楬乭拞偺悢帤偼丄堏摦帪婜偺栚埨偱偁傞丅偙偙偱丄乮徹嫆偲偟偰偼旕忢偵庛偄乯乭戞侾偺弌傾僼儕僇乭乮侾俆枩乣侾侽枩擭慜偲偝傟傞乯帪偺愨柵偟偨儂儌僄儗僋僩僗傗僱傾儞僨儖僞乕儖恖傪椺奜偲峫偊傞偲乮仏乯丄乽僲傾偺峖悈乿屻偺奼嶶偼丄乭戞俀偺弌傾僼儕僇乭丄惓妋偵偼丄乮侾夞尷傝偺乯乭弌拞搶乭偱偁傝丄俆枩乣侾侽枩擭慜偺帪婜偐傜偵側傞丅
丂丂偝傜偵丄峖悈慜偺戝婥拞偺 14俠/12俠 偺抣偑尰嵼傛傝傕偼傞偐偵彫偝偐偭偨偙偲傪峫椂偟偰廋惓偡傞偲丄尒偐偗忋偺俆乣俇枩擭慜 亖 係侽侽侽擭慜偲側傝丄乽僲傾偺峖悈乿偺帪婜偵堦抳偝偣傞偙偲偑偱偒傞丅丂偡側傢偪丄僲傾偺敔廙偐傜弌偰偒偨 僙儉丄僴儉丄儎儁僥偲偦傟偧傟偺嵢偵傛傞嵟弶偺拞搶掕廧丄偍傛傃丄偦偺屻偺僶儀儖乮乮僔僰傾儖偺抧偺乯僶儀儖偺搩 仺 悽奅傊奼嶶乯偲偄偆乭弌拞搶乭偺弌棃帠偼丄僲傾偺峖悈屻傑傕側偔偱偁傞丄偲偄偆惞彂偺婰弎偲堦抳偡傞丅乮憂悽婰侾侽丗俉丄俋乯
丂丂仏丂尰嵼偱偼丄僱傾儞僨儖僞乕儖恖偺尨巒揑側摿挜偼丄塰梴忋偺寚娮偲昦揑忬懺偵傛傞偙偲偑抦傜傟丄姰慡側僸僩偱偁傞丅乮倐倷丏僽儕僞僯僇戝昐壢帠揟丄僨儏傾儞丒俿丒僊僢僔儏丄僕僃僼儕乕丒僌僢僪儅儞摍乯丄丂僋儘儅僯儓儞恖傕姰慡側僸僩丅丂堦曽丄傾僂僗僩儔儘僺僥僋僗丄僕儍儚尨恖丄杒嫗尨恖偼丄姰慡側僒儖偱偁傝丄僸僩偲僒儖偺拞娫庬偲偄偆儈僢僔儞僌儕儞僋偼懚嵼偟側偄丅
丂